
CO2濃度が低くても気孔を開けないシロイヌナズナの変異体(patrol1変異体)を特定しました。この原因遺伝子は動物の神経伝達物質の分泌に関わるMunc13と類似したタンパクをコードしており、気孔開口に重要である事を突き止めました。PATROL1遺伝子の過剰発現体は野生株よりも大きく気孔を開くことができるため、光合成活性が上昇し、大きく成長します。この研究成果はNature Communicationsに掲載されました。
将来のCO2高濃度環境に対して植物がどのように応答するのでしょうか。その機構を分子レベルで明らかにするために、気孔のCO2応答性に異常を持つ変異体の一つ、patrol1変異体を詳しく調べました。
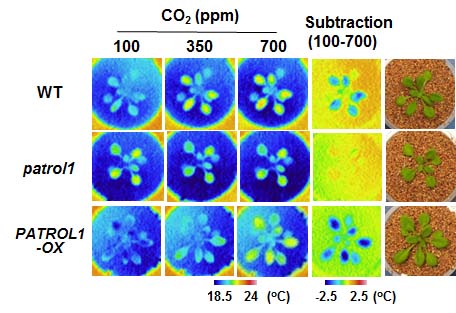
まず、多くのシロイヌナズナの中からCO2応答性の突然変異体を探し出しました。シロイヌナズナの野生株ではCO2濃度が低いほど気孔が開口し、蒸散による気化熱が奪われるため葉温は低下します。一方でpatrol1変異体はCO2濃度が低くても気孔を開けないため、野生株と比べると蒸散による気化熱の違いがありました。つまりサーモグラフィーを用いたスクリーニング法により、相対的に高温に見える株を探す事でpatrol1変異体を特定できました(図1)。
PATROL1の原因遺伝子を特定し解析した結果、MUNドメインと呼ばれる特殊なドメインを持つタンパク質をコードしていました。このドメインは動物の神経伝達物質の分泌に関わるMunc13タンパク質に存在しており、その機能は細胞内の小胞を細胞膜へ結合させ、物質の分泌や細胞膜へのタンパクの供給を行う事です。
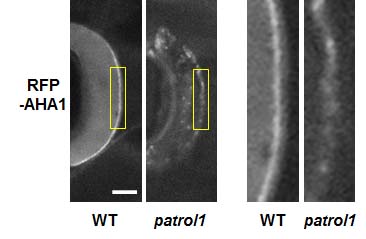
なぜpatrol1変異体は気孔を開けなくなったのかを考えてみましょう。気孔開口は細胞膜上のプロトンATPaseと呼ばれる輸送体が活性化し、プロトン(H+)放出による膜の電位差発生が引き金となっています。そこでプロトンATPaseの一つであるAHA1の細胞内での分布を調べてみると、通常は細胞膜にしか存在しないAHA1が、patrol1変異体では細胞内に点状に散在しており細胞膜上のAHA1も少なくなっていました(図2)。つまり、PATROL1の働きは、AHA1を細胞膜へ正しく局在化させることであり、patrol1変異体ではこの機能が損なわれたため気孔を開けなくなったと考えられます。
それでは逆にPATROL1の効果が大きい場合に何が起こるのでしょうか。PATROL1過剰発現体(PATROL1-OX)を調べてみると、低CO2条件では光による気孔開口が野生株よりも速く大きく起こることが明らかとなりました。さらに、光合成速度もわずかに上昇していることがわかりました。そこで、植物の成長を調べたところ、短日(夜よりも昼が短い条件)で7週間育てた植物において地上部の重量が、野生株の3割程度増加していることが明らかとなりました(図3)。

PATROL1と非常に似た遺伝子がイネ、ソルガム、ブドウなどの作物やポプラなどの樹木にも存在していますので、PATROL1遺伝子をうまく活用することでこれらのバイオマス増産に寄与できるのではないかと期待できます。
原因遺伝子同定のためのマッピングを行うとき、原因遺伝子の候補が13個ありました。中でもずば抜けて遺伝子が長く、機能が全く未知の遺伝子が含まれており、これじゃなければいいなと思っていたらこれが原因遺伝子でした。動物の方でもこれと似た遺伝子の機能は最近わかり始めたばかりで情報が少なかったです。運が悪かったと思っていたけれど、共同研究者のおかげでいい形にまとめることができ、本当に有難く思っています。
このページで使用している図はNature Communicationsからの引用となります。
より詳しく知りたい方は・・・